
Evolution des Résistances de Bio-Agresseurs au Cours de leur Expansion
Animateur: Frédéric Grognard Projet COMORE, INRIA Sophia Antipolis
Contexte général
La mise en place de plantes que nous appelerons "toxiques", c'est à dire émettant une toxine (naturelle liée à des propriétés d'antibioses ou issue de manipulation génétique) léthale pour les insectes ravageurs, fait partie des techniques de lutte qui se généralisent dans les agro-écosystèmes et leur durabilité est au coeur de la problématique de ce projet (Betz 2000). En effet, différentes variétés de maïs ou de coton exprimant des toxines issues des souches de la bactérie Bacillus thuringiensis (Bt) sont déployées dans différentes régions du monde afin de contrôler des lepidoptères; par ailleurs, des variétés de blés sont spécialement sélectionnées pour leur toxine naturelle s'attaquant à une mouche du blé. Ces relations insectes/plantes toxiques sont basées sur des interactions "gene for gene" : lorsqu'un gène de la plante est modifié, un gène de l'insecte est altéré. Ainsi, la population d'un insecte ravageur diploïde peut être classifiée en trois sous-populations : les individus susceptibles (vulnérables à la toxicité de la culture, dont les deux allèles de leur génotype seront notés SS), les individus hétérozygotes (RS) et les individus résistants (RR). Afin de limiter le développement des ravageurs résistants (RR), la mise en place de zones refuges est préconisée par des instances internationales comme l'United States Environemnental Protection Agency (EPA). Ces zones ne présentent pas de plantes toxiques et permettent donc la production d'individus susceptibles qui vont rencontrer les individus résistants émergeant des cultures toxiques. Des études théoriques (Comins 1977) montrent qu'il est possible de retarder la sélection des individus résistants si les gènes de résistance sont récessifs (un individu résistant rencontrant un individu susceptible auront une descendance susceptible). Dans ce cadre de récessivité de la résistance, la stratégie de "high dose/refuge" (HDR) a été développée pour maintenir une population de susceptibles ; cette méthode pré-suppose que la fréquence du gène de résistance dans la population est faible et que le niveau de toxine exprimé par les plantes est trés élevé (afin d'éradiquer le maximum d'individus hétérozygotes RS). Cette stratégie est préconisée pour les cultures Bt notamment (Alstad 1995). Des résistances pour certain de ces ravageurs (foreur du maïs, chenilles, mouche du blé) ont pourtant été observées au champ et la question de l'identification des mécanismes responsables de cette émergence devient prégnante. Plusieurs hypothèses ont été émises : la toxine n'est pas exprimée de manière suffisamment importante, les zones refuges sont sous-dimensionnées (Vacher et al 2003) ou la résistance est dominante (Mohammed-Awel et al 2007). C'est l'influence de la répartition des zones refuges, au niveau spatial et temporel, ainsi que les phénomènes de goulots d'étranglements sur l'évolution des résistances qui sera au coeur de notre étude. Ainsi la génétique et la dynamique des populations, les phénomènes de "bottleneck", l'hétérogénéité du milieu, les modes de dispersion à courte ou longue distance, voire les deux appelée "dispersion stratifiée" seront les élements clés pris en compte pour mener à bien ce projet.
Objectifs & Méthodo
Ce projet sera axé sur la modélisation,à base d'équations différentielles (ED) déterministes, de la génétique et la dynamique des populations répondantà des stratégies de "high dose/refuge" couplée à des phénomènes "bottleneck" d'une part et à différents processus dispersifs d'autre part.
Plus précisément, le coeur de ce travail sera la construction de modèles basés sur une approche démo-
génétique qui prend en compte la structuration de la population en fonction de ses caractéristiques
génétiques. Ce type de modèle est connu depuis les années 1930 (Kostitzin, 1936) et connait un nouvel
essor depuis peu pour décrire plus précisément les mécanismes de résistances des ravageurs (Tyutynov
et al 2008). Ils permettent de décrire l'évolution d'un génotype dont le déterminisme génétique est
assez simple, ce qui est généralement le cas pour les insectes ravageurs diploïdes, évoqués plus haut.
Il suffit en effet de considérer trois sous-populations, les susceptibles (SS), les hétérozygotes (RS), les
résistants (RR) et de décrire leurs dynamiques comme suit : 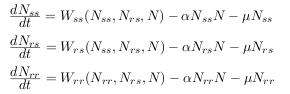 avec Nss, Nrs, Nrr les densités de populations susceptibles, hétérozygotes, résistants, alpha le taux de
rencontre et mu le taux de mortalité. Wss, Wrs, Wrr sont les fonctions de fitness et dépendent des
différents génotypes considérés. Ces fonctions de fitness sont également très fortement dépendantes de
l'environnement et changent suivant que les populations sont sur des plantes toxiques ou non.
À la différence des modèles purement populationnels, ce type de modèle permet de prendre en compte
la structuration génétique de la population et les échanges entre les sous-populations afin de décrire
de façon plus explicite les conséquences des stratégies de "high dose/refuge" sur l'évolution de ces
populations.
Dans le cadre de cette approche démo-génétique, nous nous focaliserons sur la dynamique de ces sous populations soumises d'une part à l'influence des phénomènes de goulot d'étranglements et des refuges
temporels et d'autre part à l'effet de l'hétérogéneité spatiale et des modes de dispersion des ravageurs.
avec Nss, Nrs, Nrr les densités de populations susceptibles, hétérozygotes, résistants, alpha le taux de
rencontre et mu le taux de mortalité. Wss, Wrs, Wrr sont les fonctions de fitness et dépendent des
différents génotypes considérés. Ces fonctions de fitness sont également très fortement dépendantes de
l'environnement et changent suivant que les populations sont sur des plantes toxiques ou non.
À la différence des modèles purement populationnels, ce type de modèle permet de prendre en compte
la structuration génétique de la population et les échanges entre les sous-populations afin de décrire
de façon plus explicite les conséquences des stratégies de "high dose/refuge" sur l'évolution de ces
populations.
Dans le cadre de cette approche démo-génétique, nous nous focaliserons sur la dynamique de ces sous populations soumises d'une part à l'influence des phénomènes de goulot d'étranglements et des refuges
temporels et d'autre part à l'effet de l'hétérogéneité spatiale et des modes de dispersion des ravageurs.
Bien que peu de travaux ont été effectués en ce sens, la prise en compte des phénomènes liés aux récoltes ou aux successions de plantes toxiques/non toxiques paraît essentiel. Les récoltes entrainent en effet une diminution drastique des individus dans les différentes sous-populations considérées. Les populations de faible densité résultantes peuvent alors se trouver en difficulté pour pouvoir se maintenir dans l'environnement (ce qui dépendra très fortement de leur fonction de fitness). La succession plante toxique/non toxique peut elle conduire à un goulot d'étranglement pour la population d'individus résistants. En effet, étant moins bien adaptés aux plantes non toxiques, le co^ut de la résistance peut être suffisamment important pour entrainer leur diminution voir leur disparition du milieu. Une version populationnelle a déjà été proposée en ce sens (Lemesle et al 2009) mais l'aspect demo-génétique permettrait de décrire plus précisément les interactions ravageurs/environnement. On peut notamment espèrer des préconisations plus fines sur la gestion des zones refuges.
Suivant le contexte géographique dans lequel on se situe, différents types de parcellements dépendant
de la disposition des fermes ou des exploitations agricoles peuvent être rencontrés : homogène, en
mosaïque (plus ou moins espacé).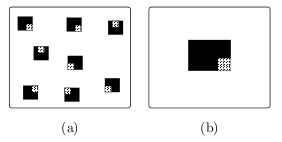 En effet, l'agriculture africaine (Afrique de l'Ouest et Centrale) est basée sur un réseau de petits
agriculteurs disséminés à l'échelle d'une région voire du pays. Cette dissémination est souvent due à la
géographie (paysages souvent accidentés, montagneux, plus ou moins fertiles) mais également au climat
trés contrasté dans ces régions tropicales (zones plus ou moins arides). Par contre, aux états-Unis, en
Australie ou encore en Inde ou en Chine, les espaces cultivés sont beaucoup plus important (Vacher 2003) : les exploitations agricoles recouvrent de très grandes surfaces et les zones refuges sont beaucoup
plus concentrées dans l'environnement. Schématiquement, l'agriculture "familiale" très développée dans
les pays du Sud peut être représentée par la figure (a) alors que les grandes exploitations mécanisées
seront plutôt du type (b). C'est l'influence de cette hétérogénéité de l'environnement sur l'évolution
des différentes sous-populations (en particulier celle des résistants) qui nous intéressera.
En effet, l'agriculture africaine (Afrique de l'Ouest et Centrale) est basée sur un réseau de petits
agriculteurs disséminés à l'échelle d'une région voire du pays. Cette dissémination est souvent due à la
géographie (paysages souvent accidentés, montagneux, plus ou moins fertiles) mais également au climat
trés contrasté dans ces régions tropicales (zones plus ou moins arides). Par contre, aux états-Unis, en
Australie ou encore en Inde ou en Chine, les espaces cultivés sont beaucoup plus important (Vacher 2003) : les exploitations agricoles recouvrent de très grandes surfaces et les zones refuges sont beaucoup
plus concentrées dans l'environnement. Schématiquement, l'agriculture "familiale" très développée dans
les pays du Sud peut être représentée par la figure (a) alors que les grandes exploitations mécanisées
seront plutôt du type (b). C'est l'influence de cette hétérogénéité de l'environnement sur l'évolution
des différentes sous-populations (en particulier celle des résistants) qui nous intéressera.
Suivant les insectes ravageurs ciblés par les plantes toxiques, leurs modes de dispersion dans l'environnement peuvent être très différents. S'ils sont polyphages leur mode de dispersion sera plutôt "continu" (c'est le cas pour certaines noctuelles, Helicoverpa armigera notamment). D'autres adoptent plutôt des stratégies de "ballooning" (individus fondateurs envoyés au hasard pour coloniser d'autre espaces, cas du puceron notamment Aphis gossypii ou encore d'insectes invasifs type Gypsy moth) : leur mode de dispersion est alors "stratifiée" (à courte et longue distance). Enfin, les ravageurs spécialistes d'une plante (qui se nourrissent presque exclusivement sur une plante, cas de certaines noctuelles comme Busseola fusca foreur du maïs) suivront eux une dispersion champ par champ (patchs à patchs).
Vers des approches couplées
Ainsi c'est la description des dynamiques temporelles d'une part ("bottleneck"/HDR) et spatio-temporelles (dispersion/HDR) d'autre part des populations de bio-agresseurs qui nous guidera tout au long de ce projet. Nous pensons que les conclusions des études précédemment citées sont très sensibles à la description des phénomènes de goulot d'étranglements, des différents types de dispersions (décrits plus haut) ainsi que du choix de modélisation de l'espace (habitat fragmenté/patchs versus homogène). Nous chercherons alors à confronter les résultats de deux types de modélisation de l'espace : métapopulations sur patchs et modèles de populations spatialement explicites. Nous nous appuierons pour cela sur les compétences complémentaires des modélisateurs participant au projet (en ED ordinaires INRIA-CIRAD, en ED partielles INRA). La participation d'entomologistes (CIRAD) est garante de la plausibilité biologique des modèles qui seront proposés et devrait permettre de les valider sur des exemples réels d'insectes ravageurs des cultures.
Sur le terrain
La finalité de ce projet est de pouvoir contribuer à la mise en évidence des paramètres clés nécessaires à la compréhension de l'évolution des résistances chez les insectes. Les expérimentations de terrain ou de laboratoire à envisager concerneront la définition des paramètres démographiques des populations. La mise en place d'élevages d'insectes des différentes sous-populations (susceptibles, résistantes ou hétérozygotes) d'une même espèce, sur milieu artificiel sera nécessaire en plus d'observations quantitatives des populations en champ. Ces observations nous permettront d'avoir accès notamment à certains paramètres des fonctions fitness Wij ; i; j = s ou r des modèles demo-génétiques développés. La capacité de dispersion des insectes devra être mesurée à l'aide des techniques apropriées (piégeages lumineux ou à base de phéromones, ou méthodes de marquage-recapture pour assurer les suivi des populations).
Références
- Kolar C.S. and Lodge D.M. 2001. Progress in Invasion Biology: Predicting Invaders. Trends Ecol. Evol. 16:199-204.
- Liebhold A.M. and Tobin P.C. 2008. Population ecology of insect invasions and their management. Annu. Rev. Entomol. 53:387-408.
- Taylor C.M. and Hastings A. 2005. Allee effects in biological invasions. Ecol. Lett. 8:895-908.
- Keitt T.H., Lewis M.A. and Holt R.D. 2001. Allee effects, invasion pinning and species' borders. Am. Nat. 157-2:203-216.
- Shigesada N. Kawasaki K. and Takeda Y. 1995. Modelling stratified diffusion in biological invasions. Am. Nat. 146-2:229-251.
- L. Mailleret and V. Lemesle. 2009. A note on semi-discrete modelling in the life sciences. Philos. T. R. Soc. A. 367:4779-4799.


